
Muscle Spindles, Proprioception, and the Golgi Apparatus
Proprioception—the sense of body position, movement and force—integrates sensory feedback from muscles, tendons, joints and the central nervous system. It underpins basic tasks like standing upright and complex skills like a gymnast’s twist or a sprinter’s stride. At the heart of this system are specialized mechanoreceptors: muscle spindles, which detect muscle length and the speed …

Proprioception—the sense of body position, movement and force—integrates sensory feedback from muscles, tendons, joints and the central nervous system. It underpins basic tasks like standing upright and complex skills like a gymnast’s twist or a sprinter’s stride. At the heart of this system are specialized mechanoreceptors: muscle spindles, which detect muscle length and the speed of stretch, and Golgi tendon organs (GTOs), which detect tension. A third player, the Golgi apparatus, operates within muscle cells as a protein‑ and lipid‑processing center. Although it does not directly sense mechanical stimuli, its role in processing and trafficking key proteins influences muscle development, metabolism and adaptation to training. Understanding these systems reveals how athletes can refine technique, enhance performance and avoid injury. Proprioceptive functions decline with aging and can be impaired by injury, but targeted training can improve body awareness and neuromuscular control. Recent research (including studies published between 2021 and 2025) has uncovered new details about spindle anatomy, gamma motor neuron modulation, proprioceptive training, and the regulation of muscle tension via the Golgi tendon organ. This article synthesizes these findings to explore how the muscle spindle, proprioception and the Golgi apparatus contribute to sports and athletic performance. It also examines training strategies that harness these systems to improve performance and reduce injury risk.
Overview of Proprioception
Proprioception is the subconscious sense of where the body and its segments are in space and how they are moving. It contributes to movement control, balance, posture, and coordination. Proprioceptive information arises from peripheral receptors—muscle spindles, Golgi tendon organs, joint receptors and cutaneous mechanoreceptors—and is integrated by the spinal cord, brainstem, cerebellum and cortex. An editorial on proprioception in sports medicine emphasised that it is critical for motor control, postural regulation and injury prevention and that integrating proprioceptive exercises into training helps athletes enhance body awareness, reduce injury risk and optimise power transfer. Moreover, proprioception has neurocognitive and emotional facets; it influences how individuals perceive their movements and can affect confidence and anxiety in athletes. Proprioceptive acuity can be improved through training. A systematic review of 19 studies found that proprioceptive training improved balance, explosive strength, speed, agility, postural stability, knee joint position sense, and technical skills; it also reduced chronic joint instability. Proprioceptive exercises include balance boards, unstable surfaces, perturbation training and dynamic movements; these challenge the sensorimotor system and encourage adaptation. Aging and sedentary behavior reduce proprioceptive function, but targeted exercises can mitigate these declines.
Muscle Spindle Anatomy and Structure
Muscle spindles are elongated sensory organs embedded within skeletal muscles. Each spindle comprises a bundle of specialized intrafusal fibers encased in a connective tissue capsule, along with afferent and efferent nerve endings and a rich capillary network. They are arranged parallel to extrafusal fibers (the contractile muscle fibers), so changes in muscle length also affect spindle length. Human spindles vary in size (up to ~8 mm long) and contain 8–20 intrafusal fibers.
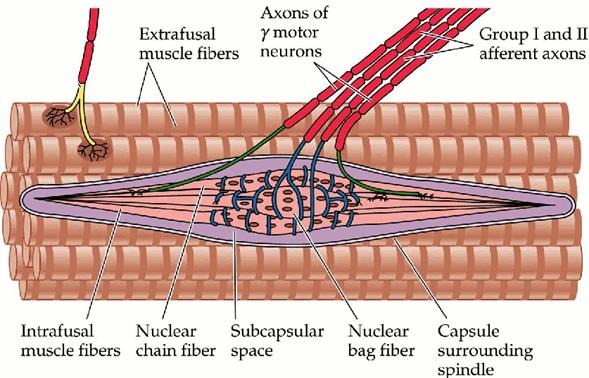
Intrafusal Fiber Types
There are three types of intrafusal fibers:
1. Nuclear bag 1 fibers (dynamic bag fibers) – thicker and longer fibers with a central region filled with nuclei; they are highly responsive to the velocity of stretch. They are innervated by type Ia afferents and by dynamic gamma motor neurons.
2. Nuclear bag 2 fibers (static bag fibers) – similar in appearance but respond more to the static length of the muscle. They receive both Ia and type II afferents and are innervated by static gamma motor neurons.
3. Nuclear chain fibers – slender fibers with nuclei arranged in a chain-like row; they respond mainly to sustained muscle length (static sensitivity) and are innervated by both Ia and II afferents and static gamma neurons.
A typical human spindle contains one dynamic bag fiber, one static bag fiber, and several chain fibers. The central region of each fiber is devoid of contractile proteins and is instead the site of sensory endings.
Sensory and Motor Innervation
Muscle spindles receive two major types of sensory (afferent) innervation:
· Type Ia (primary) afferents wrap around the central region of all intrafusal fibers in annulospiral endings. These large, fast-conducting fibers (approx. 12–20 µm diameter) signal both dynamic changes (velocity) and static muscle length. During rapid stretch, Ia afferent firing rate increases sharply.
· Type II (secondary) afferents innervate the polar regions of nuclear chain and static bag fibers through flower-spray endings. They are smaller (7–8 µm diameter) and primarily encode steady-state muscle length.
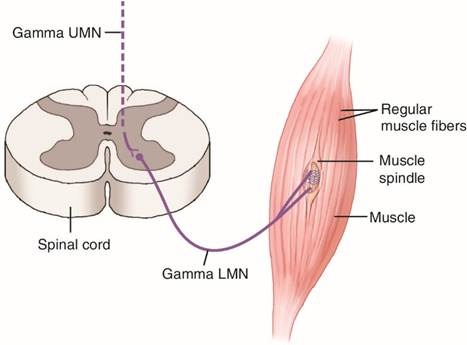
Motor innervation to the spindle is provided by gamma motor neurons (γ-motoneurons). These efferent neurons originate in the ventral horn of the spinal cord and innervate the contractile polar regions of intrafusal fibers. There are two types:
· Dynamic gamma motor neurons innervate bag 1 fibers and increase the sensitivity of spindles to changes in velocity.
· Static gamma motor neurons innervate bag 2 and chain fibers and enhance static length sensitivity.
Another class, beta motor neurons, innervates both intrafusal and extrafusal fibers but is less well understood. Gamma neurons constitute roughly 30 % of the motor pool and have smaller somata and simpler dendritic trees than α-motor neurons; they are more excitable and lack monosynaptic Ia input. Alpha-gamma coactivation ensures that muscle spindles remain sensitive during voluntary contractions. When α-motor neurons cause extrafusal fibers to contract, simultaneously activated γ-motor neurons contract the polar regions of intrafusal fibers to retension the spindle, preventing it from slackening.
Distribution and Quantity
Human muscles contain an estimated 50 000 spindles. They are present in almost all skeletal muscles except some facial muscles and tongue. Muscles requiring fine control, such as extraocular muscles and intrinsic hand muscles, have a high density of spindles, while proximal limb and postural muscles have fewer but still significant numbers. Spindles are most densely concentrated in the middle region of muscles and near the nerve entry points (NEPs) and intramuscular connective tissue; the deep layers contain more spindles than superficial layers. Trunk muscles have the highest number of spindles, whereas neck and head muscles have the highest density relative to muscle mass. Recent fascial mapping work revealed that muscle spindles are embedded within the perimysium of muscle fascial layers and maintain multiple contacts with the fascia, suggesting the state of fascial tension influences spindle sensitivity. Such mapping may help clinicians target manual therapies and stretching to modulate spindle feedback.
Muscle Spindle Physiology
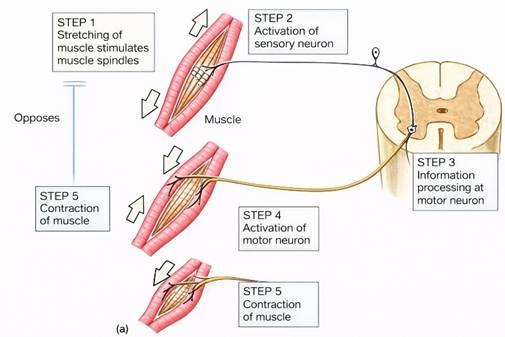
Muscle spindles transduce mechanical deformation into neural signals, providing the central nervous system with continuous information about muscle length and its rate of change. When a muscle is stretched, deformation of the spindle’s central region opens mechanically sensitive ion channels in Ia and II afferent endings, causing action potentials to propagate to the spinal cord.
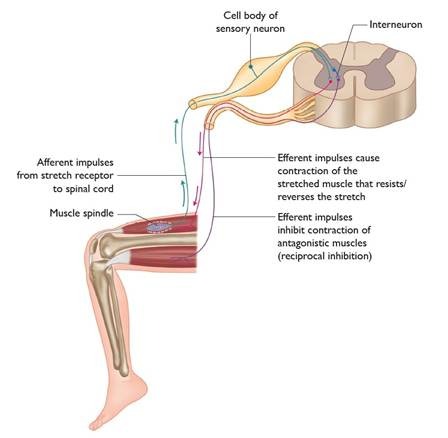
Stretch Reflex and Myotatic Response
The stretch reflex (myotatic reflex) is a monosynaptic reflex arc mediated by Ia afferents and α-motor neurons. A rapid stretch of the muscle increases Ia firing, which synapses directly onto homonymous α-motor neurons in the spinal cord, causing contraction of the same muscle. Simultaneously, Ia afferents send collateral branches to inhibitory interneurons that suppress the antagonist muscle, creating reciprocal inhibition. This reflex resists unexpected lengthening and stabilizes joints. Type II afferents also contribute to tonic muscle activity by modulating γ‑motoneuron feedback, especially during sustained postural tasks.
Dynamic and Static Sensitivity
The firing rate of Ia afferents increases in proportion to the velocity of stretch (dynamic sensitivity), whereas type II afferents primarily encode the static length. Dynamic gamma motor neurons increase dynamic sensitivity by contracting the polar ends of bag 1 fibers, making the central region tauter; static gamma neurons enhance static sensitivity by contracting bag 2 and chain fibers. The ratio of dynamic to static gamma activity varies with movement context. For precise slow movements, static sensitivity may be emphasized; for rapid or unpredictable movements, dynamic sensitivity is enhanced.
Top‑Down Control of Spindle Sensitivity
Muscle spindles are not passive sensors. The brain can tune their sensitivity via gamma and beta motor neurons. Descending pathways from the corticospinal tract, reticulospinal tract and brainstem nuclei modulate gamma activity. Research demonstrates that goal‑dependent tuning occurs during movement preparation; when planning to reach toward a target that will stretch a muscle, firing in its muscle spindles decreases before movement. This anticipatory modulation begins soon after target appearance and increases as movement onset approaches, enabling the nervous system to pre‑set muscle stiffness. Another study using mechanical perturbations found that reflex gains are strongly modulated by target direction and by the level of muscle load: reflexes become more sensitive when the muscle is unloaded and there is a long preparation time, supporting independent fusimotor control.
Adaptability and Plasticity
Spindle sensitivity is modifiable through training and mechanical manipulation. Passive ankle movement training improved proprioceptive acuity and increased the dynamic index of muscle afferent firing only when participants received explicit cues, suggesting that learning to attend to proprioceptive signals can modulate peripheral spindles. Manipulation of spindles by slackening them drastically reduced athletes’ maximal isometric adaptive force and caused earlier muscle lengthening under load. Such findings show that spindle feedback influences neuromuscular stability and can be modulated by manual techniques, with potential implications for screening and rehabilitation. Research in 2024 discovered specialized macrophages inside muscle spindles that release glutamate and directly regulate neural activity and muscle contraction. These immune cells behave like neurons and may support the metabolism of spindle fibers. The discovery highlights the interplay between the immune system and proprioception and opens new avenues for understanding fatigue and muscle disorders.
Gamma Motor Neurons and Spindle Control
Gamma motor neurons regulate the contractile state of intrafusal fibers, thereby modulating spindle sensitivity. They account for approximately one‑third of the motor neuron pool. Characteristics include small cell bodies, simpler dendritic trees and higher excitability relative to α-motor neurons. Gamma neurons lack monosynaptic Ia input and instead receive inputs from descending pathways and sensory interneurons.
Static vs. Dynamic Gamma Neurons
Static gamma neurons innervate bag 2 and chain fibers and increase baseline tension, enhancing the spindles’ response to static muscle length. Dynamic gamma neurons innervate bag 1 fibers and amplify responses to velocity changes, increasing dynamic sensitivity. In cats, static and dynamic gamma neurons can be independently controlled; humans also exhibit some independence, but the separation is less pronounced.
Alpha–Gamma Coactivation
During voluntary movement, α- and γ-motor neurons are coactivated: α-motor neurons cause extrafusal muscle contraction, while γ-motor neurons contract intrafusal fibers to keep the spindle taut. This ensures that spindles continue to signal changes in muscle length even when the muscle shortens. Without gamma activation, spindles would slacken and cease firing during contraction, leaving the nervous system “blind” to muscle length changes. Coactivation patterns can vary; for example, dynamic gamma activity increases during rapid ballistic movements, whereas static gamma activity increases during slow or postural tasks.
Stretch‑Shortening Cycle: Spindles and GTO Interplay
Many athletic movements (sprinting, jumping, throwing) involve a stretch–shortening cycle (SSC) in which a rapid eccentric contraction (muscle lengthening) is immediately followed by a concentric contraction (shortening). During the eccentric phase, muscle spindles detect rapid lengthening and elicit a stretch reflex that increases motor unit recruitment and firing rate, enhancing subsequent force production. Golgi tendon organs, conversely, inhibit muscle contraction when tension becomes excessive. For untrained individuals, this protective reflex reduces muscle stiffness and limits performance. Plyometric training reduces GTO inhibition (disinhibition), increases muscular pre‑activity and tendon stiffness, and improves performance in SSC tasks. Effective plyometrics therefore exploit spindle excitation while attenuating GTO inhibition. Warm‑up routines have similar effects: they increase motor unit recruitment, activate muscle spindles and reduce inhibition from the Golgi apparatus, contributing to post‑activation potentiation and improved power output.
Proprioceptive Training and Sports Performance
Proprioceptive training involves exercises that challenge balance, coordination and body awareness. The systematic review of 19 studies reported numerous benefits for athletes, including improved physiological capacity, balance, explosive strength, speed, agility, postural stability, knee joint position sense, muscle activation, reduction in chronic joint instability and enhanced technical ball‑control skills. Training also reduces injury risk by improving joint stability and neuromuscular control.
Types of Proprioceptive Exercises
· Balance and stability training: Using wobble boards, Bosu balls, single‑leg stance and perturbation devices to challenge postural control.
· Dynamic and reactive drills: Incorporating hops, leaps, agility ladder drills and unpredictable changes of direction to enhance dynamic joint stability.
· Unstable surface training: Performing movements on foam pads or Swiss balls to increase reliance on proprioceptive feedback.
· Sensory deprivation: Closing the eyes or reducing visual input to emphasize somatosensory feedback.
· Dual-task training: Combining cognitive tasks with balance exercises to mimic game situations and challenge attentional resources.
Evidence for Performance Enhancement
The review noted that proprioceptive training improved explosive strength, speed and agility—qualities essential for sprinting and change‑of‑direction sports. It enhanced postural stability, which benefits gymnastics, skiing and surfing. Improved joint position sense may facilitate precise shooting or throwing. Importantly, proprioceptive training reduced chronic ankle instability and associated injury rates.
Practical Recommendations
1. Integrate into warm‑ups: Include balance and coordination exercises after general warm‑up activities to prime sensory systems.
2. Progress complexity: Start with simple balance tasks and progress to dynamic and multi‑planar movements.
3. Combine with strength training: For example, perform single‑leg squats on an unstable surface to train proprioception and strength concurrently.
4. Use sport‑specific tasks: Simulate game situations where athletes must react to external stimuli while maintaining stability.
5. Consistency and variety: Regular practice is required for neural adaptations; varying tasks prevents plateauing.
Proprioceptive training should complement, not replace, traditional strength and conditioning; it works best when integrated into a comprehensive program.
Adaptations of Proprioceptors to Training
Muscle Spindle Adaptations
Research shows that muscle spindles can adapt to training:
· Passive training: A study in 2022 found that passive ankle movement training improved proprioceptive acuity only in participants who received cues. The training increased the dynamic index of muscle afferent firing, implying that spindles become more sensitive to stretch when the individual attends to proprioceptive feedback.
· Goal‑dependent tuning: Preparatory modulation of spindle firing during reaching tasks indicates that the nervous system can tune spindle sensitivity based on task goals.
· Reflex modifiability: A 2023 study demonstrated that both short‑ and long‑latency stretch reflex gains are modulated by target direction and preparation time, particularly when the muscle is unloaded. This suggests that training to anticipate movement direction can modify reflex responses via gamma control.
· Manual manipulation: Temporarily slackening spindles using manual techniques reduced athletes’ holding capacity and neuromuscular stability. This highlights the importance of proper spindle tension for force production and suggests caution when applying certain therapeutic modalities.
· Immune modulation: The discovery of glutamate‑secreting macrophages inside spindles suggests that immune cells contribute to spindle function and metabolic support. Training may influence these immune cells, although research is needed.
Golgi Tendon Organ Adaptations
Training can modify GTO responsiveness:
· Plyometric training reduces GTO inhibition, allowing greater muscle activation during explosive movements. Reduced inhibition enhances tendon stiffness and pre‑activity, leading to improved performance.
· Warm‑ups and post‑activation potentiation increase motor unit recruitment, fire spindles and reduce GTO-mediated inhibition.
· Static stretching for extended durations increases GTO firing, inhibiting spindles and enabling greater muscle lengthening. Athletes should balance stretching and strength to avoid reducing force output before performance.
Golgi Tendon Organ: Structure and Function
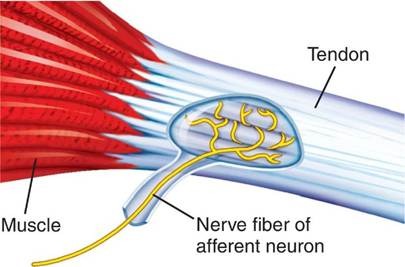
The Golgi tendon organ (GTO) is a proprioceptor located in the tendon near the musculotendinous junction. It consists of a capsule containing collagen fibers interwoven with the unmyelinated endings of a single type Ib afferent axon. Each GTO monitors tension generated by approximately 10–20 muscle fibers.
Sensory Encoding of Tension
GTOs lie in series with muscle fibers, so they are stretched whenever the muscle contracts or is passively stretched. As tension increases, the collagen fibers compress the afferent endings, generating receptor potentials that lead to action potentials in the Ib afferent. These afferents are fast‑conducting (72–120 m/s) and project to interneurons in laminae V–VII of the spinal cord. From there, signals ascend via the dorsal columns to the cerebral cortex, contributing to conscious perception of force and position.
Autogenic Inhibition and Reflexes
When tension exceeds a set threshold, GTO afferents activate inhibitory interneurons that suppress the activity of α-motor neurons innervating the same muscle, causing muscle relaxation—a phenomenon known as autogenic inhibition. This inverse myotatic reflex prevents muscle damage during excessive force production. The reflex is complemented by excitatory effects on antagonist muscles, promoting coordinated movement. Prolonged static stretching (low-force stretch held for >7 seconds) activates GTOs, reduces muscle spindle firing and increases muscle length tolerance.
Role in Postural Stability and Movement
GTOs are particularly dense in lower-limb extensor muscles; they encode the level of load and contribute to postural adjustments during standing and locomotion. The interplay between GTO and spindle signals allows the nervous system to coordinate muscle stiffness and force production. In the context of explosive movements, such as jumping, the inhibitory effect of GTOs can limit performance; however, training can modulate this response.
Cellular Golgi Apparatus: Structure and Function
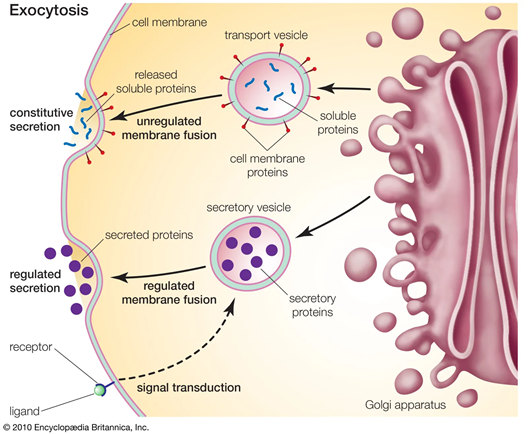
Although the Golgi apparatus is not a proprioceptor, it plays a crucial role in muscle cell function and adaptation. Discovered by Camillo Golgi in 1898, the organelle consists of stacked flattened membrane sacs (cisternae) organized into cis, medial and trans compartments, collectively forming a ribbon-like structure held together by a matrix of scaffold proteins. Newly synthesized proteins and lipids from the endoplasmic reticulum (ER) enter the cis-Golgi, undergo sequential modifications (glycosylation, sulfation, phosphorylation) as they pass through the medial cisternae, and are sorted and packaged in the trans-Golgi network for delivery to lysosomes, secretory granules or the plasma membrane. The Golgi apparatus also participates in lipid synthesis and transport, signal transduction, and cell-cycle regulation. Its structural integrity is critical; fragmentation or dispersal disrupts protein processing and is associated with diseases such as muscular dystrophy, neurodegeneration and metabolic disorders. In muscle cells, the Golgi apparatus is dispersed into numerous elements rather than forming a single ribbon as in typical secretory cells.
The Golgi Apparatus in Skeletal Muscle
Distribution and Fiber‑Type Differences
Skeletal muscle fibers have thousands of small Golgi elements distributed throughout the sarcoplasm. A pioneering study using rat muscle showed that slow‑twitch (type I) fibers have most Golgi elements near the plasma membrane, while fast‑twitch (type IIB) fibers have more internal Golgi elements. After denervation, Golgi distribution shifts toward the slow‑twitch pattern, suggesting neural activity influences Golgi organization. Subsequent work demonstrated that patterned electrical stimulation of denervated muscle influences Golgi distribution: fast stimulation maintains fast-fiber characteristics with fewer surface elements, whereas slow stimulation increases Golgi density at the fiber surface and organizes microtubules accordingly. These findings indicate that neural activity and muscle usage patterns regulate Golgi positioning and microtubule organization.
Functional Significance
The dispersed Golgi elements in muscle likely serve specialized roles, such as:
· Glycosylation of membrane proteins and receptors: Ion channels, transporter proteins (e.g., glucose transporter GLUT4) and signaling receptors require proper glycosylation for trafficking and function. Golgi elements near the sarcolemma may process proteins destined for the membrane.
· Lipid metabolism: Exercise-induced increases in the enzyme sphingomyelin synthase 1 (SGMS1), which localizes to the Golgi apparatus, produce diacylglycerols (DAGs) used to synthesize phospholipids. These lipids are transported to mitochondria for cardiolipin and phosphatidylethanolamine synthesis, improving mitochondrial function and insulin sensitivity. Thus, exercise may remodel the Golgi lipid metabolism to enhance muscle health.
· Autophagy and protein quality control: Muscle fibers must repair damaged proteins and organelles after intense exercise. The Golgi, along with the ER, contributes to the formation of autophagosomes and the sorting of lysosomal enzymes.
Implications for Sports Performance
While research on the Golgi apparatus in athletics is limited, the findings above suggest that neural activation patterns and training intensity could influence Golgi organization, protein processing and lipid metabolism. For instance, fast power training may favour fast‑fiber Golgi distribution patterns (internal elements), whereas endurance training may promote surface-associated Golgi and enhanced lipid processing. Aberrant Golgi function might contribute to muscle fatigue or metabolic disorders, whereas targeted training could support healthy Golgi activity.
Integration of Proprioception and Golgi Apparatus with Athletic Performance
Muscle spindles, Golgi tendon organs and the Golgi apparatus operate at different levels of biological organization but together influence athletic performance.
1. Sensorimotor Integration: Spindles and GTOs provide real‑time feedback on muscle length and tension, enabling precise control of movement. Their interplay during the stretch–shortening cycle underlies explosive actions like jumping, sprinting and throwing. Proper tuning of spindle sensitivity via gamma motor neurons allows athletes to adjust muscle stiffness to meet task demands, while modulating GTO inhibition through plyometric training maximizes force output.
2. Neuromuscular Adaptation: Proprioceptive training enhances reflex sensitivity, balance and coordination, leading to improved performance and reduced injury risk. Training modifies spindle and GTO responses, possibly through changes in gamma motor neuron excitability and central processing. Warm‑ups and post‑activation potentiation prime proprioceptors and reduce Golgi-mediated inhibition, boosting subsequent power performance.
3. Cellular and Metabolic Adaptation: At the cellular level, the Golgi apparatus processes and traffics proteins and lipids critical for muscle function. Neural activity patterns regulate its organization; exercise alters its lipid metabolism (via SGMS1) and may influence glucose transport and mitochondrial health. Efficient Golgi function ensures that muscle fibers can synthesize and transport receptors, ion channels and structural proteins essential for contraction and recovery.
4. Holistic Perspective: Understanding the combined roles of proprioceptors and cellular organelles offers a holistic perspective on athletic performance. Training programs should consider how sensory feedback, neural control and cellular metabolism interact to produce movement. For instance, athletes recovering from injury might incorporate proprioceptive exercises to restore spindle sensitivity while nutritional strategies support Golgi-mediated protein and lipid processing.
Conclusion and Future Directions
Muscle spindles, proprioception and the Golgi apparatus are fundamental to human movement and athletic performance. Spindles detect muscle length and velocity through specialized intrafusal fibers and are tuned by gamma motor neurons. They mediate stretch reflexes, inform the brain about body position, and adapt to training. Golgi tendon organs monitor tension and protect muscles via autogenic inhibition but can inhibit performance when overactive; training techniques such as plyometrics can reduce this inhibition. The Golgi apparatus within muscle cells processes and traffics proteins and lipids and adapts to neural activity patterns, affecting muscle metabolism and health. Athletes and coaches should incorporate proprioceptive training into programs to enhance balance, agility and injury resistance. Warm‑ups and plyometric exercises can optimize spindle responsiveness and modulate GTO inhibition. Nutrition and recovery strategies should support healthy Golgi function, ensuring efficient protein processing and lipid metabolism. Future research should explore how different training modalities influence spindle and Golgi adaptations, how immune cells within spindles contribute to neuromuscular function, and how metabolic interventions can enhance performance.
By integrating insights from neurophysiology and cell biology, we can better understand the intricate systems that enable human movement. This knowledge not only informs athletic training but also aids rehabilitation and supports lifelong mobility.